
This brief account summarizes some basic features of marine invertebrate larvae, with remarks on their phylogenetic significance. It is intended for non-specialists or students new to the subject. The remarkable diversity of form among marine larvae was first appreciated in the mid-19th century, when they were discovered in plankton by Johannes Müller and his students. The larvae illustrate the tendency of evolution to produce variations on the theme in an especially vivid fashion, and as such, they appear prominently in the writings of evolutionary biologists during the latter half of the century. We are still uncertain as to the precise evolutionary relationship between larvae of different phyla. In recent years, major contributions to our understanding of larval diversity and evolution have come especially from Scandinavian biologists, notably Gosta Jägersten, Gunnar Thorson and, most recently, Claus Nielsen (see references).
According to current thinking, the diversity of form among marine larvae largely reflects the specific adaptations required for planktonic feeding and locomotion, but this occurs against a background of evolutionary constraints that arise from the inherited ground plan and developmental sequence. It remains a major task to separate these two components, so that specific and largely secondary specializations can be distinguished from the basic features on which a phylogenetic sequence can eventually be built.
Top of Page
Not all invertebrate phyla with marine representatives have pelagic larvae. Many, including those specialized for small body size, e.g. most ecdysozoans, have direct development, whereby the hatching stage is simply a small version of the adult. Among the invertebrates that have larvae, there is usuaully a well-defined type larva for each phylum. Each type is presumably primitive larval type for the phylum, and similarities between them can be usd in some instances to link them in a phylogenetic series. How far back in time such series can be taken is a matter of debate. Jägersten, at one extreme, has postulated an ancestral pelagobenthic life cycle (pelagic larva + benthic adult) for metazoans as a group, which would makes pelagic larvae very ancient indeed. Larvae descended directly from the ancestral type, assumed to resemble the planula of modern cnidarians, are referred to as primary larvae. The majority of pelagic marine invertebrate larvae (i.e. those considered below) fall into this category. A number of secondarily evolved larvae (secondary larvae) also occur among invertebrates. These are generally modified juvenile stages of what were originally direct-developing groups, and are seen most notably in arthropods, parasitic flatworms and chordates.
Much of the interest in invertebrate larvae stems from the assumption that the primary larva concept is correct to at least a degree. This would mean that a good deal of phylogenetically useful information can probably be extracted from modern larvae by judicious application of morphological and molecular techniques, and this is currently an active area of investigation.
Top of Page
The most useful generalization to emerge from modern studies is the recognition that primary marine larvae can be separated into two basic types, depending on whether they are organized in a downstream or an upstream fashion. The terms refer to the position of the main ciliary band and the direction of ciliary beat in relation to the mouth. In downstream larvae, the mouth is located behind (posterior to) the principal band, whose cilia beat towards it. This means that locomotory and feeding currents coincide. The larvae can thus feed as they swim, and are typically rather swimmers with a compact body.
In upstream larvae, in contrast, the mouth is anterior to the main locomotory ciliary bands, and the latter beat away from the mouth. Water currents generated during locomotion cannot then be used directly for food capture. Instead, a variety of indirect means of concentrating food particles are employed. These appear to depend largely on hydrodynamic effects generated by the complex topology of the body surface [specific ciliary behaviors, e.g. reversals, may also be involved in some instances]. In consequence, upstream larvae are often large, with large surface areas, swim slowly, and have surprisingly bizarre shapes. They are also usually transparent and rather delicate, in part because rapid growth to comparatively large size, using minimal amounts of tissue, evidently confers a significant adaptive advantage. Upstram larvae consequently make excellent microscopical subjects, and it is no accident that they figure more prominently than downstream larvae in research on the mechanics of development whenever direct observations on live larvae are required.
Phylogenetic trees constructed on the basis of larval type accord well with those based on other criteria, e.g. molecular sequences. Two major, well-defined groups emerge, protostomes and deuterostomes, the former with downstream larvae and the latter with upstream ones. Larval morphology thus provides additional support for the premise that the protostome/deuterostome distinction is a natural one of long evolutionary standing.
Top of Page
 |
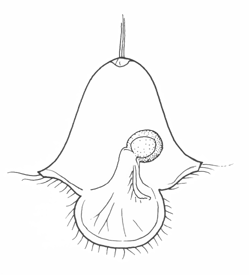 |
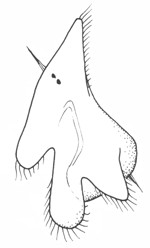
The simplest of downstream larvae are Müller's larva (and the smaller Götte's larva) of polyclad flatworms and the pilidium of nemertines. These are topologically the same: both have an anteriorly positioned apical organ or apical tuft, a ventroposterior mouth leading to a blind gut with no anus, and a circumferential ciliary band or set of bands. Polyclad larvae are are generally compact, solid and opaque. The band runs along the margins of a set of projecting lobes, six or eight in number. As the larva matures, the body enlarges axially at the expense of the lobes, which shrink and gradually disappear.
The pilidium is helmet-shaped, with lateral lobes like ear-flaps on each side. The band runs along the margins of the helmet, and the larva itself is delicate and transparent. The juvenile develops internally in a remarkable process involving the ingrowth of a number of imaginal discs, which apply themselves to the stomach and expand and fuse to enclose it. The juvenile worm then emerges through the larval epithelium and, over a period of minutes, consumes the latter as its first meal.
Among the more advanced marine protostomes, trochophore larvae are characteristic of those traditionally classed as spiralians. This includes annelids, sipunculids, echiurids and some mollusks (e.g. chitons), though most other mollusks pass through a provisional trochophore stage. The trochophore, or something very much like it, is thus very probably a basal larva for spiralians.
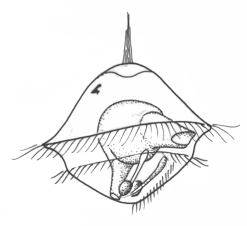 |
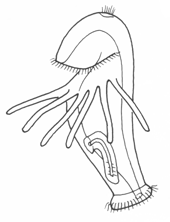
Trochophores are named for the wheel-like appearance of the main ciliary band (the prototroch), which girdles the body around its middle. The mouth opens just below the prototroch, and the anus is posterior and terminal. Beside the latter are a pair of nephridia and the mesoblast cells that later give rise to the trunk rudiment. The apex is occupied by an apical tuft, variously modified in different groups; the juvenile cerebral ganglia develop on either side of the tuft. Metamorphosis can be rapid, but is not usually dramatic. It typically involves the loss of the trochal band and adjacent tissue, which reshapes the head, and the expansion of the segmented trunk. Some trochophores (e.g. serpulids, echiurids and Polygordius, and archiannelid) have an accessory feeding band, the metatroch, just below and parallel to the prototroch. The prototroch and metatroch cilia beat in reverse direction, and food particles are propelled along the groove between them (the food groove) to the mouth. This arrangement is referred to as an opposed-band feeding system. Other trochophores have evolved alternative feeding mechanisms, often involving fleshy lips or specialized oral ciliature. It is not clear whether the metatroch is a primitive feature of basal spiralians or a later specialization, but this is a potentially interesting topic for future research.
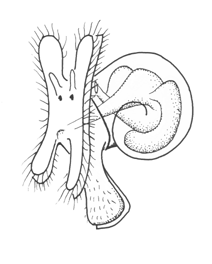 |
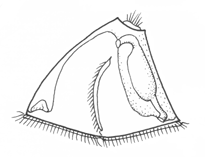
Among mollusks, non-feeding larvae, some trochophore-like, others less so, are found among the aplacophorans and chitons. Gastropods and bivalves have veliger larvae, which differ from each other somewhat because of the different shell form. The figure shows the gastropod veliger. It has the basic topology of a trochophore, but with an expanded ventral foot region with an operculum, a shell secreted to enclose the visceral mass, and an expanded pair of velar lobes with a food groove running along its edge. The expanded velar lobes greatly lengthen the surface available for food collection, but the arrangement is otherwise essentially the same as in trochophores with a metatroch, and the molluscan velum is presumably derived from this.
The sequence below shows, in schematic form, the relation between the various spiralian larvae, beginning with a hypothetical planula-type ancestor with a posterior mouth and cilia. Whether or not these were ever organized as a separate band in the ancestral larva, as shown, the direction of beat is always towards the mouth. Moving the mouth forward along the side of the body, as in Müller's larva or the pilidium, would then define an effective ventral surface. Adding an anus and metatroch then gives us, in sequence, the trochophore and veliger.
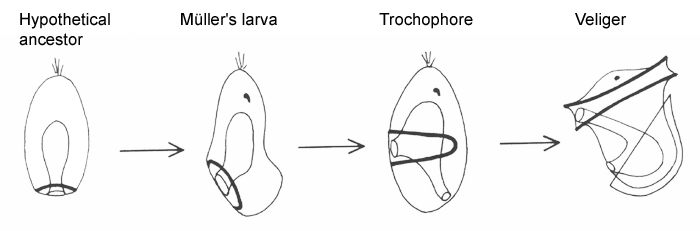 |
 |
 |
Lophophorates (phoronids, brachiopods and bryozoans) are puzzling for combining features of both protostomes and deuterostomes. In terms of larval organization, this is seen in the presence of a preoral hood with cilia that beat towards the mouth, like a prototroch, while bands posterior to the mouth, typically arrayed on tentacles or ridges, beat away from the mouth. Current molecular data allies the lophophorates with spiralians (together they form the Lophotrochophora), which suggests the postoral band is probably a derived feature, perhaps precociously developed from the tentacles of the adult at some time in the past. There are numerous examples of this kind of heterochronic borrowing among marine larvae; the process is known as adultation. Two lophophorate larvae are illustrated here, the actinotroch (phoronids) and the cyphonautes (bryozoans). Their common organization is evident in that either is easily converted into the other, simply be expanding or reducing the preoral hood as shown in the following sequence.
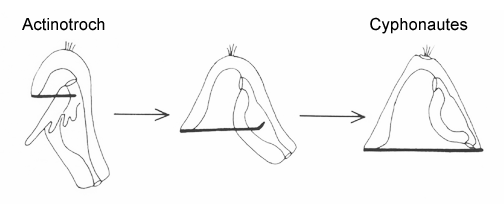 |
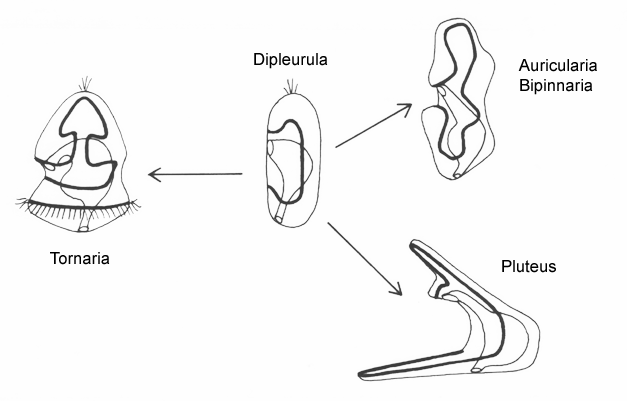
The planktonic feeding larvae of lower deuterostomes (echinoderms + hemichordates) are all variants of a generalized type known as a dipleurula. The principal band in dipleurula larvae encircles the mouth, and the cilia beat away from it. The zone between the band and mouth (the oral field) can also be ciliated, and these cilia typically beat towards the mouth. The mouth is thus not upstream of all the body cilia, only the majority, including those in the band.
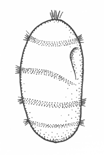
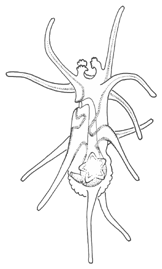
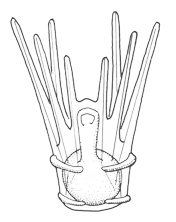
There are two basic strategies to increase the efficiency of food collection in upstream dipleurula-type larvae. In the tornaria of enteropneust hemichordates, the starfish (asteroid) bipinnaria and the sea cucumber (holothurian) auricularia, the band becomes convoluted as the larva grows, so as to enclose a complex and precisely contoured oral field. As the larvae swim, food particles are directed towards the mouth, at least in part by the hydrodynamic effect of the contouring. A second solution, seen in pluteus larvae, is to extend the band along a set of projecting arms supported by skeletal rods. The larvae swim with the arms projecting forward, and food is swept towards the mouth. By analogy, the larvae acts much like a catcher's mitt pushing forward into a field of food particles.
 |
 |
 |
Of the larvae that rely on contouring, the auricularia is unusual for the dramatic fragmentation and reorganization of the ciliary bands that takes place at the onset of metamorphosis. These convert the auricularia into a doliolaria, a barrel-shaped stage with a series of 4 or 5 circumferential bands. Crinoids have a doliolaria as well, and lack feeding larvae altogether. It is not clear which type of larva, feeding or non-feeding, is ancestral for the phylum as a whole. The bipinnaria differs from the auricularia in developing a series of projecting lobes. These are muscular and flex, providing an alternative means of locomotion for the advanced-stage larvae, which are too large for ciliary propulsion to be very effective. In addition, as the bipinnaria matures, a three short brachiolar arms develop near the apex. These are used for attachment while the juvenile rudiment develops. The larvae at this stage is known as a brachiolaria.
Tornaria larvae enhance their swimming capabilities by a different means: they develop a posterior accessory band with compound cilia, the telotroch, that provides the main propulsive force. Metamorphosis is less catastrophic than in echinoderms; the tornaria is converted gradually on settlement into a vermiform enteropneust.
 |
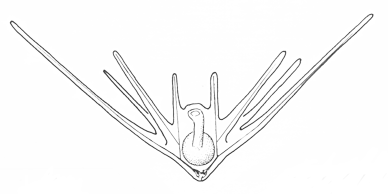 |
Pluteus larvae, which occur in echinoids (sea urchins) and ophiuroids (brittle stars) rely on an internal skeleton of calcarious spicles for support and body form. The two pluteus types are superficially similar in overall appearance, but are clearly convergent in this respect and independently evolved from something more like the bipinnaria. Evidence for this is seen in the posterior-most of the larval arms, which are differently positioned in echinoid and ophiuroid plutei, and hence are not homologous. There are also differences in swimming efficiency. Echinoid plutei are comparatively small, and are fast swimmers; ophiuroid are weak swimmers and use their arms more as flotation devices. Plutei can generally be identified on the basis of the skelatal structure, which is varied and species specific.
Top of Page
The larvae of the invertebrate chordates (tunicates and amphioxus) are secondary larvae whose relation to the primary dipleurula-type larvae of echinoderms and hemichordates is somewhat obscure. Past authorities have argued that chordates actually derive from motile larvae that have evolved to become sexually mature (paedomorphosis). One such proposal, more specific in its predictions than most, is due to Walter Garstang: he noted the topological similarity between dipleurula ciliary bands (which have their own innervation) and chordate neural plate, and suggested that the latter actually derive from the former. While there is as yet no convincing evidence that chordates are directly derived from the body of a dipleurula-type larva, the core of the idea, that there is continuity in the neurogenic tissues, has some appeal. It is a surprisingly parsimonious way of deriving a characteristic chordate structure, the neural tube, whose origin is otherwise unexplained.
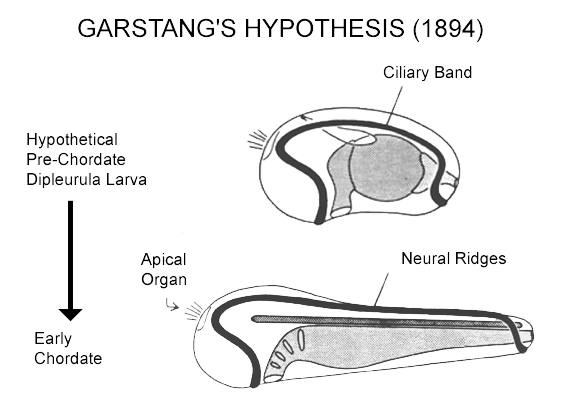 |
There are two problems with Garstang's hypothesis, however, both a consequence of comparatively recent data on the expression patterns of the genes required to pattern the body during embryonic development. First, dipleurula-type larvae fail to express a full set of antero-posterior patterning genes. Instead, they mainly express genes associated with the head region, which makes them less than ideal models for an ancestral form for chordates, which do clearly have a full set of patterning genes. More specifically, it seems unlikely that the whole length of the nerve cord in chordates could have arisen from a ciliary band of an ancestral larva that has no equivalent of the chordate trunk and tail.
The second problem is that chordates appear to be dorsoventrally inverted relative to other metazoans, including those (i.e. echinoderms and hemichordates) with dipleurula larvae. Note that with Garstang's idea, the dorsal body surface always remains dorsal through the transition to chordates. A variant of Garstang's hypothesis that avoids this problem has been proposed the Nielsen (1999). He accommodates the inversion problem by having the brain originate from a post-oral site on the original ventral side of the body, which, after inversion, becomes dorsal. The mouth is then also dorsal, but this is the case also in some basal chordates, namely tunicates. Secondarily the mouth would have to move to a ventral position in more advanced chordates, i.e. amphioxus and vertebrates. At first sight, a post-oral origin for the brain is very counterintuitive, but the idea is increasingly well supported by the emerging molecular data."
Testing the validity of this and related ideas, in order to better understand the link between lower deuterostomes and protochordates, is an active area of current research in evolutionary biology.
Top of Page