
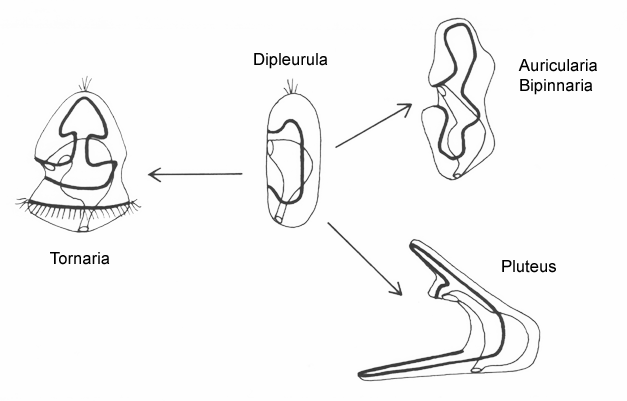
The planktonic feeding larvae of lower deuterostomes (echinoderms + hemichordates) are all variants of a generalized type known as a dipleurula. The principal band in dipleurula larvae encircles the mouth, and the cilia beat away from it. The zone between the band and mouth (the oral field) can also be ciliated, and these cilia typically beat towards the mouth. The mouth is thus not upstream of all the body cilia, only the majority, including those in the band.
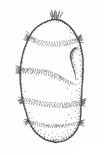
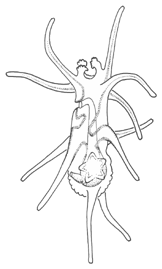
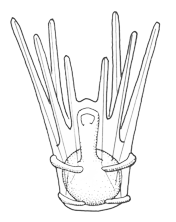
There are two basic strategies to increase the efficiency of food collection in upstream dipleurula-type larvae. In the tornaria of enteropneust hemichordates, the starfish (asteroid) bipinnaria and the sea cucumber (holothurian) auricularia, the band becomes convoluted as the larva grows, so as to enclose a complex and precisely contoured oral field. As the larvae swim, food particles are directed towards the mouth, at least in part by the hydrodynamic effect of the contouring. A second solution, seen in pluteus larvae, is to extend the band along a set of projecting arms supported by skeletal rods. The larvae swim with the arms projecting forward, and food is swept towards the mouth. By analogy, the larvae acts much like a catcher's mitt pushing forward into a field of food particles.
 |
 |
 |
Of the larvae that rely on contouring, the auricularia is unusual for the dramatic fragmentation and reorganization of the ciliary bands that takes place at the onset of metamorphosis. These convert the auricularia into a doliolaria, a barrel-shaped stage with a series of 4 or 5 circumferential bands. Crinoids have a doliolaria as well, and lack feeding larvae altogether. It is not clear which type of larva, feeding or non-feeding, is ancestral for the phylum as a whole. The bipinnaria differs from the auricularia in developing a series of projecting lobes. These are muscular and flex, providing an alternative means of locomotion for the advanced-stage larvae, which are too large for ciliary propulsion to be very effective. In addition, as the bipinnaria matures, a three short brachiolar arms develop near the apex. These are used for attachment while the juvenile rudiment develops. The larvae at this stage is known as a brachiolaria.
Tornaria larvae enhance their swimming capabilities by a different means: they develop a posterior accessory band with compound cilia, the telotroch, that provides the main propulsive force. Metamorphosis is less catastrophic than in echinoderms; the tornaria is converted gradually on settlement into a vermiform enteropneust.
 |
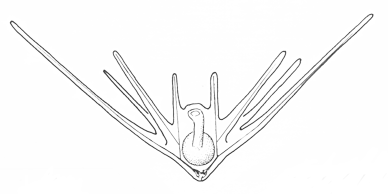 |
Pluteus larvae, which occur in echinoids (sea urchins) and ophiuroids (brittle stars) rely on an internal skeleton of calcarious spicles for support and body form. The two pluteus types are superficially similar in overall appearance, but are clearly convergent in this respect and independently evolved from something more like the bipinnaria. Evidence for this is seen in the posterior-most of the larval arms, which are differently positioned in echinoid and ophiuroid plutei, and hence are not homologous. There are also differences in swimming efficiency. Echinoid plutei are comparatively small, and are fast swimmers; ophiuroid are weak swimmers and use their arms more as flotation devices. Plutei can generally be identified on the basis of the skelatal structure, which is varied and species specific.